| Fowell, S. J., Cornet, B., and Olsen, P. E., 1994, Geologically rapid Late Triassic extinctions: Palynological evidence from the Newark Supergroup, in Klein, G. D., ed., Pangea: Paleoclimate, Tectonics, and Sedimentation During Accretion, Zenith, and Breakup of a Supercontinent: Boulder, Colorado, Geological Society of America Special Paper 288. |
Geologically rapid Late Triassic extinctions: Palynological evidence from the Newark Supergroup
S.J. Fowell, B. Cornet*, and P.E. Olsen
Department of Geological Sciences and
Lamont-Doherty Earth Observatory of Columbia University,
Palisades, New York 10964
ABSTRACT Orbitally controlled, sedimentary cycles of the Newark Supergroup permit palyniferous Late Triassic sections to be calibrated in time. Carnian palynofloras from the Richmond basin exhibit 2-m.y. fluctuations in the spore/pollen ratio, but taxonomic composition remains stable. Diversity of Norian and Rhaetian palynofloras increases prior to a 60% reduction at the Triassic/Jurassic boundary. The extinction of Late Triassic palynomorph species is coincident with a spike in the spore/pollen ratio and approximately synchronous with the last appearances of tetrapod taxa and ichnofossil genera. This geologically brief episode of biotic turnover is consistent with bolide impact hypotheses. |
*Present address: 27 Tower Hill Avenue, Red Bank, New Jersey 07701
INTRODUCTION
The composition and diversity of Triassic faunas and flo- ras are of particular interest with respect to the late Norian (=Rhaetian) mass extinction, an event that culminated in a 42% decrease in the number of terrestrial tetrapod families (Olsen et al., 1987) and a 23% reduction in diversity of marine invertebrate families (Sepkoski, 1984). Although the Norian event is recognized as one of the most severe Phanerozoic mass extinction episodes, substantial disagreement remains as to the duration and synchroneity of the Late Triassic terrestrial and marine extinctions.Multiple extinction events throughout the Carnian and Norian have been proposed for both terrestrial tetrapods (Benton, 1986) and marine invertebrates (Benton, 1986; Johnson and Simms, 1989). Hallam (1981) argued that the extinctions were concentrated in the latter part of the Norian (=Rhaetian), but he maintained that the tetrapod turnover preceded extinction of the marine invertebrates by several million years. Olsen et al. (1987) provided evidence that Triassic vertebrate faunas persisted until the latest Triassic.
Although these hypotheses make different predictions about the Late Triassic fossil record, the dearth of Triassic/Jurassic boundary sections (Hallam, 1981), the difficulty of correlating marine and terrestrial strata, and the proliferation of Late Triassic time scales (see Kerp, 1991) render them difficult to test.
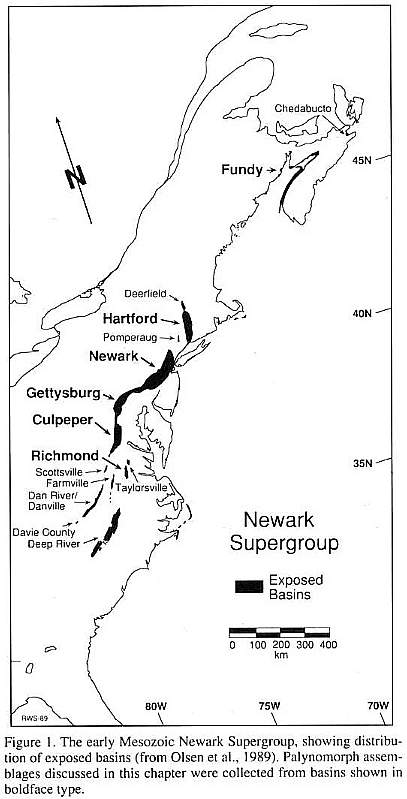
These problems can be partially surmounted in the Newark Supergroup of eastern North America (Fig. 1). Sedimentation in these rift basins was continuous from the Carnian through the Hettangian, and the Triassic/Jurassic boundary is accessible in five separate basins. The presence of dated basalt horizons, in conjunction with periodic sedimentary cycles, per- mits construction of a Late Triassic time scale calibrated in absolute time (Fig. 2) (Fowell and Olsen, 1993). No marine strata are present, but the variety of fossilized materials, including osseous tetrapod remains, palynomorphs, and vertebrate ichnofossils, permits comparison of a suite of organisms preserved under diverse conditions. In this chapter, we deal primarily with the palynological record. However, it is hoped that the numerous tetrapod fossils and palynomorph-bearing horizons will ultimately enable more rigorous correlations with global terrestrial and marine strata.
CYCLICITYThe Newark Supergroup is composed predominantly of fluvial and lacustrine strata that fill a series of rift basins formed by the Pangean breakup. During the Late Triassic and Early Jurassic, this chain of elongate half grabens stretched from 4�S to 15�N (Witte et aI., 1991). Climate models for idealized geographic representations of Pangea have shown that a large landmass centered on the equator will exhibit summer and winter monsoon circulation patterns (Kutzbach and Gallimore, 1989; Short et al., 1991). These models are consistent with the thick lacustrine deposits of the Newark Supergroup, where long-term fluctuations of annual precipitation are indicated by the presence of hierarchical transgressive and regressive lacustrine cycles (Olsen, 1986; Olsen et al, 1989).
The shortest of these cycles are shallowing-upward packages between 1.5 and 35 m thick, identified in outcrop on the basis of rock type and fabric (Olsen, 1986; Olsen et al., 1989; Olsen, 1991). Using the rationale outlined by Olsen (1986), cycles have been calibrated via biostratigraphic age estimates and published radiometric dates. Biostratigraphic calibration and Fourier analysis of these cycles both yield an average periodicity of approximately 21 k.y. (Olsen, 1986). This value falls between the 18-k.y. and 21.5-k.y. precession periods calculated for the Early Jurassic (Berger et al., 1992).
Fourier analysis of long stratigraphic columns and cores of the entire Triassic Newark basin section reveals higher-order cycles with periodicities of 100 k.y., 400 k.y., and 2 m.y. (Olsen, 1986). The l00 k.y. and 400 k.y. cycles correspond to periodicities predicted by Milankovitch theory for the Earth's eccentricity cycles (Hays et al., 1976; Short et al., 1991), which have remained relatively constant throughout Earth history (Berger and Pestiaux, 1984). A 2-m.y. eccentricity cycle can also be shown to exist from the orbital dynamics, but unlike the other cycles its effect on climate has not been modeled.
The regularity and duration of the Newark Supergroup lacustrine cycles are compatible with the hypothesis that orbitally controlled seasonal and latitudinal variations in solar radiation detennined the length and intensity of Mesozoic rainy seasons. A latitudinal climatic gradient is also apparent; differences in the type and abundance of sedimentary structures and facies indicate that southern Newark Supergroup basins were far more humid than the northern basins (Hubert and Mertz, 1980; Olsen, 1991; Olsen et al., 1989). This drier- to-the-north trend resulted in three primary types of lacustrine cycles distinguishable by the ratio of wet/dry fabrics: the humid Richmond type, the dry Fundy type, and the intermediate Newark type (Olsen, 1991). Palynologically productive horizons are generally restricted to the deeper-water lacustrine facies. Consequently, the completeness of the palynological record in stratigraphic sections from the Newark Supergroup increases with the average lake depth.
Dry lacustrine cycles occur in the Fundy basin of Nova Scotia (Fig. I), where sedimentation rates were so low that 21- k.y. cycles cannot be identified. Higher-order cycles are manifested in the fonn of sand-patch cycles (Smoot and Olsen, 1985, 1988). These cycles are approximately 1 to 2 m thick and consist of sedimentary fabrics that record alterations between shallow, perennial lakes and playas with salt crusts (Olsen, 1991). Deep-water lacustrine facies are extremely rare, and preserved palynofloras are similarly sparse.
Intermediate Newark-type cycles are present in the Dan River/Danville, Culpeper, Gettysburg, Newark, Pomperaug, Hartford, and Deerfield basins (Fig. 1). These 21-k.y. cycles display a range of sedimentary fabrics, from deep-water mi- crolaminae to subaerial desiccation cracks. Many of the cycles contain deep-water facies, but shallow-water and desiccated fabrics are more common (Olsen, 1991). Palynomorph preservation is restricted to the deep-water facies and is discontinuous within individuaI 21-k.y. cycles.
Humid Richmond-type cycles are present in the Rich mond and Taylorsville basins of the southern Newark Supergroup (Fig. 1). Desiccated, mudcracked fabrics are rare, and 21-k.y. cycles consist primarily of microlaminated claystones, coals, and shallow-water siltstones (Olsen, 1991). The nearly continuous palynological record of the Richmond basin is ideal for studies of early Late Triassic microfloral diversity.
PALYNOFLORAL ASSEMBLAGES
AND SEDIMENTARY CYCLICITY
OF THE CARNIAN RICHMOND BASIN
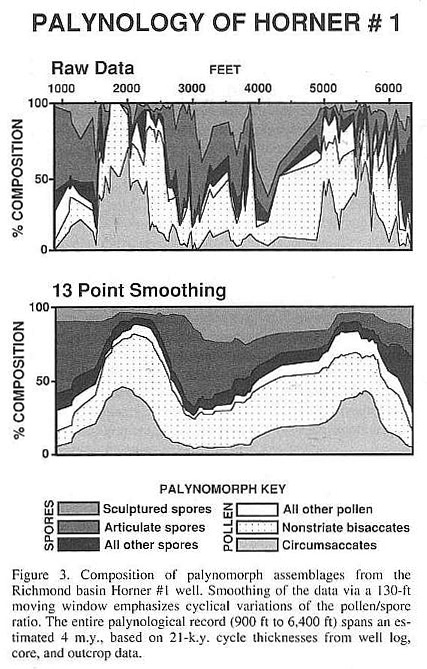
Outcrops are rare in the Richmond basin, but a nearly complete palynological record has been obtained from the 2,000-m-deep Horner #1 well (Fig. 3). Well cuttings of the entire middle Carnian Richmond basin section, from the Otterdale Formation to the base of the Tuckahoe Formation, were collected at 9.1-m (30-ft) intervals and processed for palynomorphs..
In contrast to Early Jurassic palynomorph assemblages, which are dominated by a single pollen genus (Corollina), the morphological and generic diversity of the Richmond basin palynofloras is characteristic of Triassic assemblages through- out the Newark Supergroup. Dominance of either bisaccate or monosaccate (circumsaccate) pollen morphotypes is also typical. Spores, however, are far more abundant in the Carnian Richmond basin assemblages than in Norian palynofloras from northern basins. The Horner #1 well cuttings exhibit large fluctuations in the relative abundance of pollen and spores (Fig. 3) that appear to reflect the 2-m.y. climate cycle.
Well logs from Horner #1 indicate that the thickness of the 21-k.y. cycle increases from 7 to 10 m downsection. Adjacent to the Horner #1 well, a 150-m outcrop of the Turkey Branch Formation also exhibits an extremely regular, 7-m cycle thickness. Cores of underlying formations confirm the presence of thicker, 10-m cycles. The average 21-k.y. cycle thickness is estimated to be 8.5 m for the palynologically productive portions of the Horner well. Application of this estimate to the data in Figure 3 demonstrates that the Horner #1 well spans approximately 4 m.y. Peaks of the pollen/spore ratio, emphasized by smoothing of the data in Figure 3, occur at ~2-m.y. intervals. The spores were produced by lower vascular plants that favored high precipitation and/or humidity, whereas pollen-producing seed plants dominated during intervals of relatively arid climate.
Despite periodic fluctuations in palynomorph abundances, the taxonomic composition of the assemblages remains relatively stable throughout the entire Carnian record of the Richmond basin. This indicates that the climatic variations produced gradual floral changes and were not of sufficient magnitude to cause regional extinctions.
LATE TRIASSIC AND EARLY JURASSIC PALYNOFLORAS
The youngest sediments in the Richmond basin are of Carnian age, but the palynological record is continued through Norian, Rhaetian, and Hettangian strata in basins to the north. The Triassic/Jurassic boundary in these basins has been identified by the appearance of palynofloras dominated by the circumpolloid pollen genus Corollina. This Early Jurassic Corollina meyeriana palynoflora (Fig. 2) rapidly replaces the diverse, monosaccate- and bisaccate-dominated assemblages characteristic of the Carnian, Norian, and Rhaetian (Cornet, 1977; Cornet and Olsen, 1985).
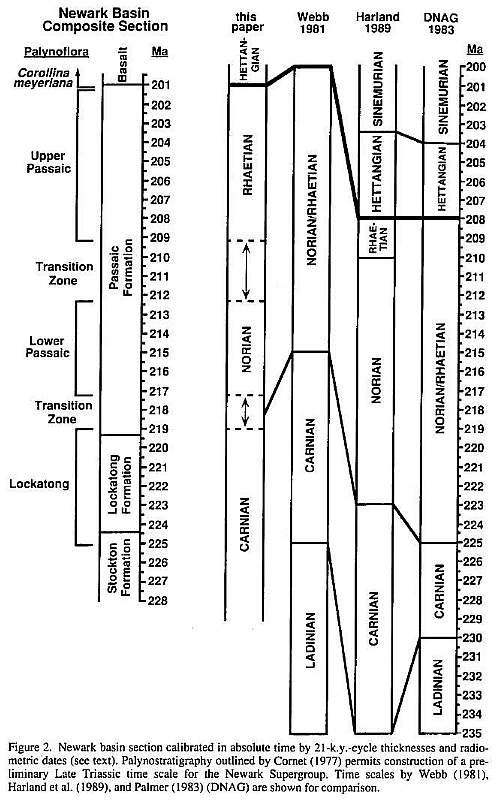
The abrupt transition from Late Triassic to Early Jurassic palynofloras is evident in Figure 4, a composite range chart of palynomorph species from deep-water, lacustrine facies of the Culpeper, Gettysburg, Hartford, and Newark basins (see Table 1 for species names with citations). Temporal resolution of these ranges is derived from Newark basin cyclostratigraphy. Fowell and Olsen (1993) use measured cycle thicknesses from the Late Triassic and earliest Jurassic lacustrine record of the Newark basin to calibrate the stratigraphic section in relative time (Fig. 2). This section is fixed in absolute time by 202.2 � 1.3 Ma 40 Ar/39 Ar dates (Sutter, 1988) and 201 � 1 Ma U/Pb dates (Dunning and Hodych, 1990) of the Palisades sill, which fed the Orange Mountain basalt and the correlative Jacksonwald basalt (Ratcliffe, 1988).
The composite range chart in Figure 4 was constructed on the time-calibrated Newark basin section using the graphic correlation method outlined by Shaw (1964). As discontinuous preservation precluded calculation of continuous correlation lines from the palynological data alone, correlation lines were fixed by paleontological, stratigraphic, and palynological tie points (Fowell and Olsen, 1993).
Species' ranges for the late Carnian, Norian, and Rhaetian (Fig. 4; Table 1) indicate that palynofloral diversity increased throughout the last 24 m.y. of the Triassic (see Fig. 5 of Fowell and Olsen, 1993). The transition from diverse Triassic assemblages to Corollina-dominated Jurassic palynofloras occurs at approximately 201 Ma and is coincident with a loss of 13 of the 20 most common Late Triassic species. In total, 24 of the 40 species recorded from the latest Triassic palynofloras are absent from Early Jurassic assemblages. This represents a 60% regional extinction across the Triassic/Jurassic boundary.
TRIASSIC/JURASSIC BOUNDARY SECTIONS
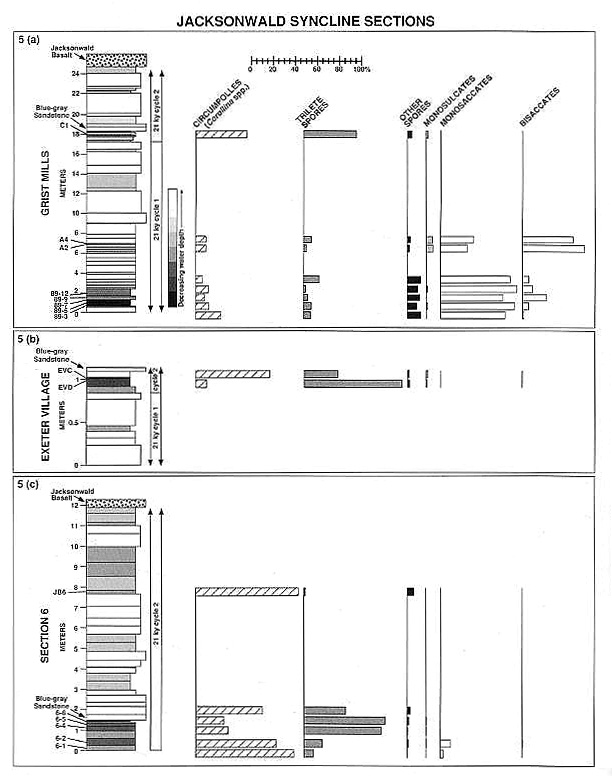
Palynomorph assemblages from exposures in the Jacksonwald Syncline of the Newark basin further limit the Triassic/Jurassic boundary palynoflora! turnover to an 11-m interval that is barren of palynomorphs. A local cycle thickness of ~20 m allows the duration of this transition to be constrained to less than 21 k.y. (Fig. 5) (Fowell and Olsen, 1993).
Palynomorph assemblages characterized by unusually high percentages (>50%) of trilete spores have been recovered from the Triassic/Jurassic boundary in three Jacksonwald syncline sections (Fig. 5a, b, and c). All three sections lie within the Passaic Formation, and two extend to the base of the ~201 Ma Jacksonwald basalt. Stratigraphic correlations are facilitated by the cyclostratigraphy of the sections and the presence in each of a black, organic-rich clay layer overlain by an unusual, blue-gray, plant-bearing sandstone. Spore-dominated assemblages Cl, EVD, 6-4, and 6-5 are composed of 50 to 89% trilete spore species of the genera Anapiculatisporites, Converrucosisporites, Deltoidospora, Dictyophyllidites, Granulatisporites, Kyrtomisporis, Porcellispora, Reticulatisporites, Todisporites, and Verrucosisporites (R. Litwin, personal communication, 1989; Olsen et al., 1990). These palynofloras occur between the highest Triassic assemblage (A4, Fig. 5a) and the lowest Jurassic Corollina meyeriana palynoflora (EVC, Fig. 5b).
Palynofloras from section 6 (Fig. 5c) and the Exeter Village section (Fig. 5b) serve to bracket the stratigraphic duration of the spore-dominated assemblages. The percentage of trilete spores increases markedly between samples 6-2 and 6-4 of section 6 and decreases again from 6-5 to 6-6. The abundance of trilete spores also drops off dramatically above sample EVD of the Exeter Village section.
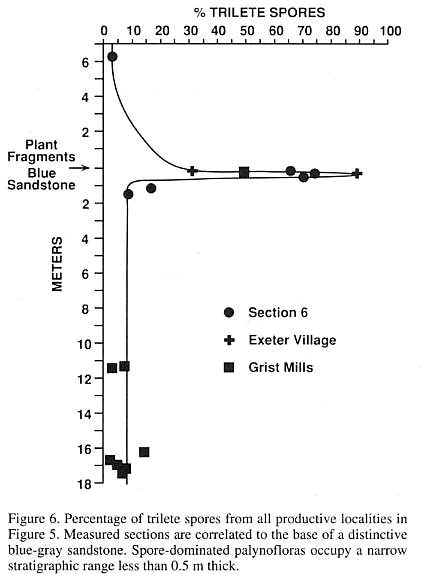
In Figure 6, spore percentages from all productive localities in the Jacksonwald Syncline are plotted against thickness above and below the base of the blue-gray sandstone. The spore-dominated palynofloras have a limited stratigraphic distribution (less than 0.5 m), apparently originating and terminating abruptly.
In the Grist Mills (Fig. 5a) and Exeter Village (Fig. 5b) sections, this spore spike occurs above typical Late Triassic assemblages of monosaccates and bisaccates and below Corollina-rich Jurassic palynofloras. This relationship is not apparent in section 6 (Fig. 5c). Samples 6-1 and 6-2, which underline the spore spike, contain the high percentages of Corollina typical of Jurassic assemblages. Yet these assemblages retain low abundances of the monosaccate Triassic index species Vallasporites ignacii and Patinasporites densus. The only comparable palynofloras in the Newark Supergroup are from the Fundy basin, where samples with abundant Corollina and rare (but clearly not reworked) Patinasporites densus are present 32 cm below the North Mountain basalt. This basalt has been assigned a U/Pb date of 202 � 1 Ma (Hodych and Dunning, 1992) and is correlative with the Jacksonwald basalt. Given the low sedimentation rates in the Fundy basin (Olsen et al., 1989; Olsen, 1991), the Fundy assemblages may be time-equivalent with samples 6-1 and 6-2. We consider these palynofloras indicative of uppermost Triassic strata due to the presence of diagnostic Triassic species, which are absent from overlying assemblages in both the Newark and Fundy basins.
DISCUSSION AND CONCLUSIONS
Evidence for a geologically rapid, terminal-Triassic biotic turnover in the Newark Supergroup is not confined to the palynological record. Footprint assemblages from the Jacksonwald Syncline show that the last appearances of the Late Triassic ichnofossil genera Gwyneddichnium, Apatopus. Brachychirotherium. and two new genera are coincident with the palynologically defined Triassic/Jurassic boundary (Silvestri, 1991; Silvestri and Szajna, 1993). Absence of these five footprint taxa from Early Jurassic assemblages corresponds to the disappearance of previously abundant tetrapod trackmakers and constitutes a decrease of approximately 50% in the diversity of Newark basin ichnofossil genera (Silvestri, 1991; Silvestri and Szajna, 1993).




Terrestrial tetrapod fossils of Late Triassic age are preserved throughout the Newark Supergroup. However, no characteristic Triassic taxa are found among abundant osseous remains from the earliest Jurassic strata of the Fundy basin. Calibration of the ranges of the Late Triassic vertebrate families by lacustrine cycles indicates that the last appearances of dominant Triassic taxa occurred within an interval of 850 k.y. that brackets the Triassic/Jurassic boundary (Olsen et al., 1987).
The abruptness and apparent synchroneity of the tetrapod, ichnofossil, and palynofloral extinctions in the Newark Supergroup are consistent with catastrophic extinction scenarios involving volcanism or bolide impacts. Fowell and Olsen (1993) previously rejected volcanism as a catalyst for Late Triassic palynological turnover in the Newark Supergroup on the grounds that the first basalt flows lie 7 to 10 m above the transition from Triassic to Jurassic palynomorph assemblages and ash-fall horizons have not been observed.
Bice et al. (1992) report the discovery of multiple shocked quartz horizons from the Triassic/Jurassic boundary in Tuscany, Italy; the uppermost horizon coincides with the abrupt disappearances of Rhaetian bivalves and Late Triassic foraminifera. Bice et al. (1992) suggest that the presence of three shocked quartz layers fits the comet shower model proposed by Hut et al. (1987).
Aspects of the Newark Supergroup palynofloral record are consistent with a multiple-impact scenario. The presence of a geologically brief spore-spike above the last appearances of Late 'Triassic palynomorph species is analogous to Cretaceous-Tertiary boundary fern spikes (Tschudy et al., 1984; Tschudy and Tschudy, 1986; Nichols et al., 1986; Fleming, 1990) hypothesized to represent a recolonization of ferns following catastrophic destruction of the Cretaceous flora.
The presence of short-lived "transitional" assemblages, in which Corollina is dominant but rare Late Triassic species persist (samples 6-1 and 6-2, Fig. 5c), indicates possible disruption of the flora prior to the Triassic/Jurassic boundary turnover. It is tempting to suggest a relationship between these palynofloras and one of the lower shocked quartz horizons of Bice et al. (1992) and to correlate the spore-spike with the uppermost shocked quartz layer and the extinctions of Triassic bivalves and foraminifera. Such correlations are premature, however, and require better estimates of sedimentation rates in the Tuscany section and evidence for shocked quartz in the Newark basin sections. To date, a comprehensive search for shocked quartz in the Newark Supergroup has not been completed, but we are currently conducting this crucial test of the validity of the impact hypothesis.
Enhanced temporal resolution and accurate stratigraphic correlations are also essential to future comparisons of Triassic/Jurassic boundary sections. Periodic sedimentary cycles and good palynofloral preservation within the Newark basin allow temporal calibration of Late Triassic palynofloral zones. Preliminary results are shown in Figure 2 and compared to published chronostratigraphic time scales. Recently acquired cores of the entire Triassic Newark basin section will permit refinement of the temporal data and better resolution of the boundaries between palynofloral zones. It is hoped that the combination of palynological and tetrapod data from the Newark Supergroup will eventually enable correlation between our continuously calibrated Late Triassic time scale and global sections with ammonite control.
ACKNOWLEDGMENTS
We thank S. Silvestri and R. Litwin for access to their unpublished data. We also thank Superior Oil Company (now Mobil) for the funds to process the Horner #1 well samples and R. Jones for accomplishing the bulk of the Horner well processing. We are grateful to A. Traverse for sharing his laboratory during the initial phases of this study and providing useful discussion. B. Coakley, A. Traverse, and an anonymous reviewer read the manuscript and suggested changes that significantly improved it. Their time and effort is greatly appreciated. This research was supported by a grant from the National Science Foundation (EAR 89 16726).
REFERENCES CITED
Benton, M, J., 1986, More than one event in the late Triassic mass extinction: Nature, v. 321, p. 857-861.
Berger, A., and Pestiaux, P., 1984, Accuracy and stability of the Quaternary terrestrial insolation, in Berger, A. L., Imbrie, J., Hays, J., Kukla, G., and Saltzman, B., eds., Milankovitch and climate: Dordrecht, Reidel, p. 83-111.
Berger, A., Loutre, M. F., and Laskar, J., 1992, Stability of the astronomical frequencies over the Earth's history for paleoclimate studies: Science, v. 225, p. 560-566.
Bice, D. M., Newton, C. R., McCauley, S., Reiners, P. W., and McRoberts, C. A., 1992, Shocked quartz at the Triassic-Jurassic boundary in Italy: Science, v. 255, p. 443-446.
Comet, B., 1977, The palynostratigraphy and age of the Newark Supergroup [Ph.D. thesis]: University Park, The Pennsylvania State University, 505 p.
Comet, B., and Olsen, P. E., 1985, A summary of the biostratigraphy of the Newark Supergroup of eastern North America with comments on early Mesozoic provinciality, in Weber, R., ed., III Congreso Latinoamericano de Paleontologia Mexico, Simposio Sobre Floras del Triasico Tardio, su Fitogeografia y Paleoecologia: Memoria: Mexico City, Universidad National Autonoma de Mexico, Instituto de Geologia, p. 67-81.
Dunning, G. R., and Hodych, J. P., 1990, U/Pb zircon and baddeleyite ages for the Palisades and Gettysburg sills of the northeastern United States: Implications for the age of the Triassic/Jurassic boundary: Geology, v. 18, p. 795-798.
Fleming, R. F., 1990, Nonmarine Cretaceous-Tertiary boundary, southwestern United States: Geological Society of America, Abstracts with Programs, v. 22: A278.
Fowell, S. J., and Olsen, P. E., 1993, Time-calibration of Triassic/Jurassic microfloral turnover, eastern North America: Tectonophysics, v. 222, p.361-369.
Hallam, A., 1981, The end-Triassic bivalve extinction event: Palaeogeography, Palaeoclimatology, Palaeoecology, v. 35, p. 1-44.
Harland, B. W., Armstrong, R. L., Cox, A. V., Craig, L. E., Smith, A. G., and Smith, D. G., 1989, A geologic time scale: Cambridge, Cambridge University Press, 263 p.
Hays, J. D., Imbrie, J., and Shackleton, N. J., 1976, Variations in the Earth's orbit: Pacemaker of the ice ages: Science, v. 194, p. 1121-1132.
Hodych, J. P., and Dunning, G. R., 1992, Did the Manicouagan impact trigger end-of-Triassic mass extinction?: Geology, v. 20, p. 51-54.
Hubert, J. F., and Mertz, K. A., 1980, Eolian dune field of Late Triassic age, Fundy Basin, Nova Scotia: Geology, v. 8, p. 516-519.
Hut, P., and seven others, 1987, Comet showers as a cause of mass extinc- tions: Nature, v. 329, p. 118-126.
Johnson, A.L.A., and Simms, M. J., 1989, The timing and cause of late Triassic marine invertebrate extinctions: Evidence from scallops and crinoids, in Donovan, S. K., ed., Mass extinctions: New York, Columbia University Fress, p. 174-194.
Kerp, H., 1991, Some recently published (numerical) time scales compared: Albertiana, v. 9, p. 19-21.
Kutzbach, J. E., and Gallimore, R. G., 1989, Pangaean climates: Mega- monsoons of the megacontinent: Journal of Geophysical Research, v. 94, p.3341-3357.
Nichols, D. J., Jarzen, D. M., Orth, C. J., and Oliver, P. Q., 1986, Palynological and iridium anomalies at Cretaceous/Tertiary boundary, south-central Saskatchewan: Science, v. 231, p. 714-717.
Olsen, P. E., 1986, A 40-million-year lake record of early Mesozoic orbital climatic forcing: Science, v. 234, p. 842-848.
Olsen, P. E., 1991, Tectonic, climatic, and biotic modulation of lacustrine ecosystems-examples from Newark Supergroup of eastern North America, in Katz, B., ed., Lacustrine basin exploration: Case studies and modem analogs: American Association of Petroleum Geologists Memoir 50, p. 209-224.
Olsen, P. E., Shubin, N. H., and Anders, M. H., ]987, New Early Jurassic tetrapod assemblages constrain Triassic-Jurassic tetrapod extinction event: Science, v. 237, p. 1025-1029.
Olsen, P. E., Schlische, R. W., and Gore, J. W., eds., 1989, Tectonic, depositional, and paleoecological history of Early Mesozoic rift basins, eastern North America (International Geological Congress field trip guidebook T351): Washington, D.C., American Geophysical Union, 174 p.
Olsen, P. E., Fowell, S. J., and Cornet, B., 1990, The Triassic/Jurassic boundary in continental rocks of eastern North America; A progress report, in Sharpton, Y. L., and Ward, P. D., eds., Global catastrophes in Earth history: Geological Society of America Special Paper 247, p. 585-593.
Palmer, A. R. (compiler), 1983, The decade of North American geology, 1983 geologic time scale: Geology, v. 11, p. 503-504.
Ratcliffe, N. M., 1988, Reinterpretation of the relationship of the western ex- tension of the Palisades sill to the lava flows at Ladentown, New York, based on new core data, in Froelich, A. J., and Robinson, G. R., Jr., eds, Studies of the Early Mesozoic basins of the eastern United States: U.S. Geological Survey Bulletin 1776, p. 113-135.
Sepkoski, J. J., Jr., 1984, A kinetic model of Phanerozoic taxonomic diversity. III: Post-Paleozoic families and mass extinctions: Paleobiology, v. 10, p.246-267.
Shaw, A. B., 1964, Time in stratigraphy. New York, McGraw Hill, 365 p. Short, D. A., Mengel, J. G. Crowley, T. J., Hyde, W. T., and North, G. R., 1991, Filtering of Milankovitch cycles by earth's geography: Quaternary Research, v. 35, p. 157-173.
Silvestri, S. M., 1991, Ichnofauna of the last 7 million years of the Triassic from the Jacksonwald Syncline, Newark Basin, Pennsylvania [Master's thesis]: Newark, New Jersey, Rutgers University, 94 p.
Silvestri, S. M., and Szajna, M. J., 1993, Biostratigraphy of vertebrate footprints in the Late Triassic section of the Newark basin: Reassessment of stratigraphic ranges, in Lucas, S. G., and Morales, M., eds., The nonmarine Triassic: New Mexico Museum of Natural History and Science Bulletin 3, p.439-445.
Smoot, J. P., and Olsen, P. E., 1985, Massive mudstones in basin analysis and paleoclimatic interpretation of the Newark Supergroup, in Robinson, G. R., Jr., and Froelich, A. J., eds., Proceedings, Second U.S. Geological Survey Workshop on the Early Mesozoic Basins of the Eastern United States: U.S. Geological Survey Circular 946, p. 29-33.
Smoot, J. P., and Olsen, P. E., 1988, Massive mudstones in basin analysis and paleoclimatic interpretation of the Newark Supergroup, in Manspeizer, W., ed.; Triassic-Jurassic rifting, continental breakup and the origin of the Atlantic Ocean and passive margins: New York, Elsevier, p. 249-274.
Sutter, J. F., 1988, Innovative approaches to dating igneous events in the early Mesozoic basins of the eastern United States, in Froelich, A. J., and Robinson, G. R., Jr., eds., Studies of the Early Mesozoic basins of the eastern United States: U.S. Geological Survey Bulletin 1776, p. ]94-200.
Tschudy, R. H., and Tschudy, B. D., ]986, Extinction and survival of plant life following the Cretaceous/Tertiary boundary event, Western Interior, North America: Geology, v. 14, p. 667-670.
Tschudy, R. H., Pillmore, C. L., Orth, C. J., Gilmore, J. S., and Knight, J. D., 1984, Disruption of the terrestrial plant ecosystem at the Cretaceous-Tertiary boundary, western interior: Science, v. 225, p. 1030-1032.
Webb, J. A., ]981, A radiometric time scale of the Triassic: Journal of the Geological Society of Australia, v. 28,. 107-121.
Witte, W. K., Kent, D. Y., and Olsen, P. E., 1991, Magnetostratigraphy and paleomagnetic poles from Late Triassic-earliest Jurassic strata of the Newark basin: Geological Society of America Bulletin, v. 103, p. 1648-1662.
MANUSCRIPT ACCEP'TED BY THE SOCIETY MAY 14, 1993Printed in U.S.A.