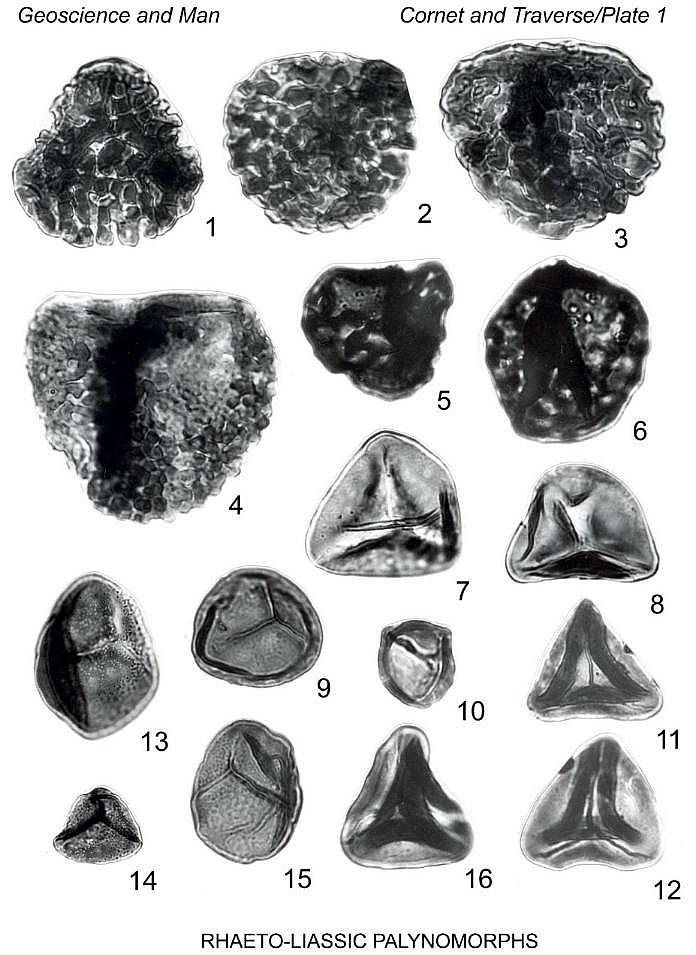
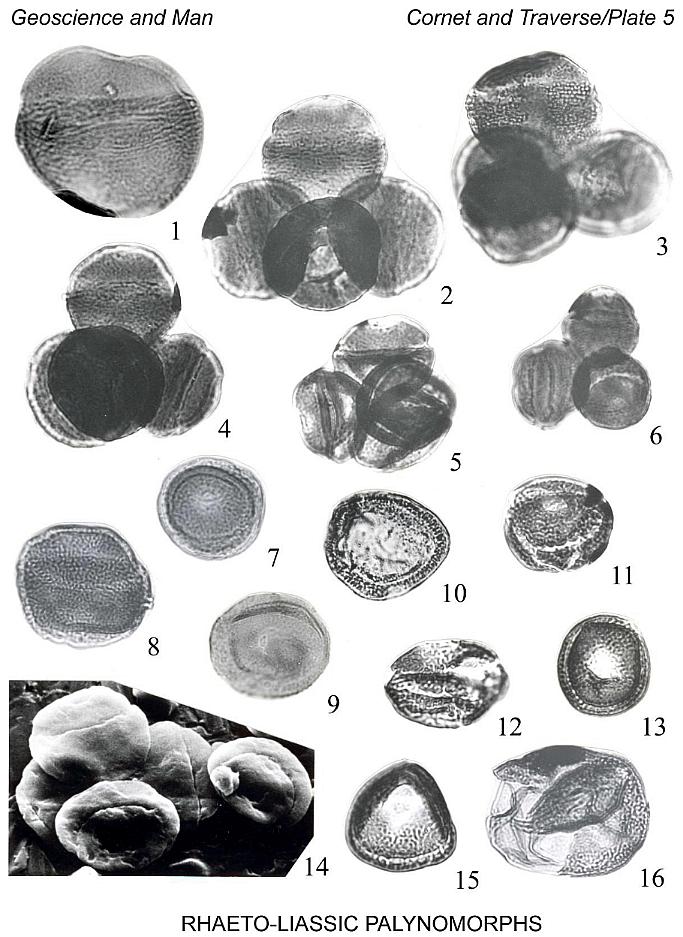
Holotype: Plate 1, fig. 13, size 46 um; Slide AGA6-1, coordinates 52.8 x
111.2, ref . 28.5 x 111.6.
Description: Miospores, trilete. Amb subspherical, rarely subtriangular.
Laesurae margins slightly thickened, sometimes as narrow conspicuous lips. Radius of
laesura 35-46% of overall diameter of spore. Exine 0.6-0.8 um thick, proximally covered
with densely to sparsely spaced pits, 0.3-0.4 um in diameter. Pits more closely spaced
near laesura, decreasing in number equatorially; distal side generally lacking pits. Size
range (11 specimens): 25-52 um (median 41um).
Specific epithet refers to Agawam, Mass.
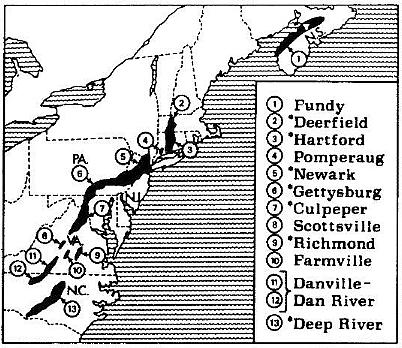
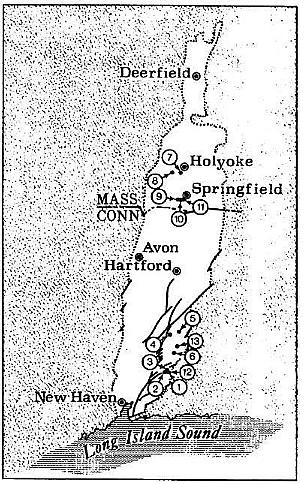
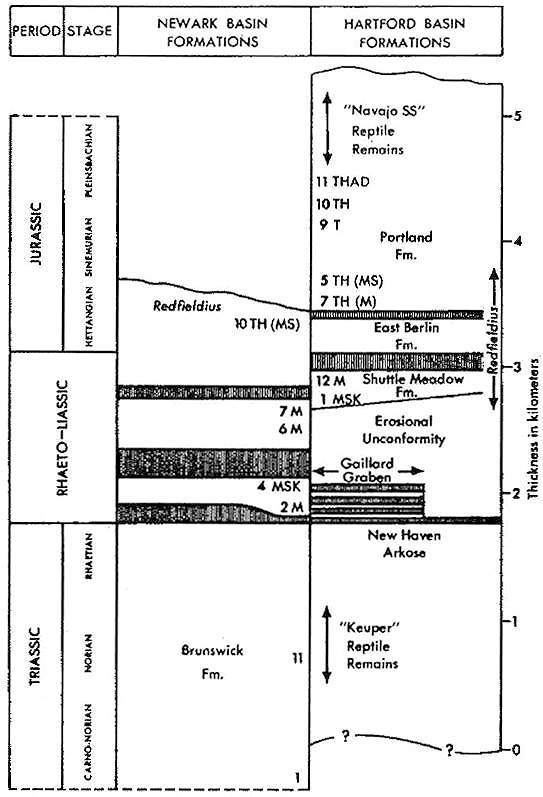
Locality: Portland Fm., 11 (rare).
Genus Gleicheniidites Ross 1949
Type species: Gleicheniidites senonicus Ross 1943
Gleicheniidites cf G. nilssonii Pocock 1970
Plate 1, figure 16
Description: See Pocock (1970, p. 32).
Remarks: The uniqueness of this species necessitates some description: our
specimens compare very closely with G. nilssonii, but have a slightly thicker exine (about
2.6 um thick between apices, decreasing to about 2.1 um at apices). A distal triangular
area of thickening is delimited by 3 arcuate ridges, intersecting at the apices. Two
specimens measure 41 um and 43 um in diameter. The small number of specimens permits only
tentative assignment at this time.
Locality: Portland Fm., 5 (rare).
Genus Granulatisporites Ibrahim 1933 emend. PotoniÕ & Kremp 1954
Type species: Granulatisporites granulatus Ibrahim 1933
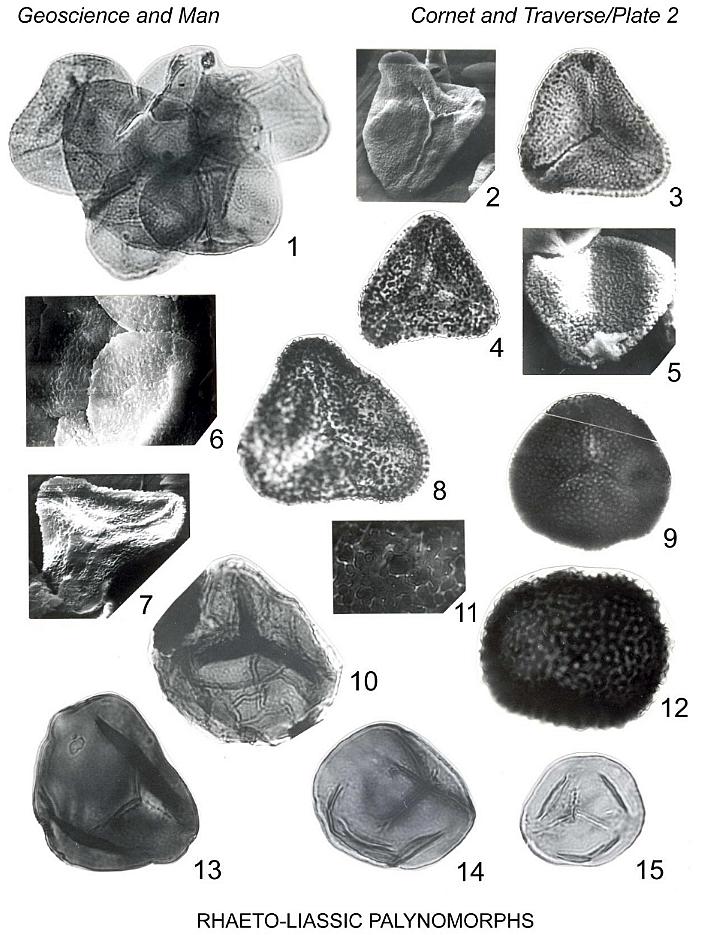
Granulatisporites infirmus (Balme 1957) comb. nov.
Plate 2, figures 1, 2
Concavisporites infirmus Balme, 1957, p. 21, Pl. 2, fig. 32, 33 (Basionym).
Trachysporites asper Nilsson, 1358, p. 39, P1. 2, fig. 3.
Granulatisporites cf. G. asper (Nilsson) Playford & Dettman, 1965,
p. 135, Pl. 12, fig. 10.
Emended description (more than 500 specimens): Isospores radial, trilete. Amb
usually subtriangular with broadly rounded apices, and sides slightly concave to slightly
convex; amb sometimes subspherical. Laesura extends almost to, rarely reaching, equatorial
margin, usually bordered by narrow, slightly elevated lips. Kyrtomes prominently to weakly
developed, not always present. Laesurae frequently bordered by arcuate folds concave
toward proximal pole. Exine sparsely to densely sculptured with grana and small verrucae,
mixed with a few bacula and coni, all of which are frequently joined at their bases by a
low, imperfect ridge system or incipient reticulum. Sculpture is comprehensive, but
usually proximally scattered and more reduced; sculpture ranges from predominantly small
grana and verrucae, 0.1-0.8 um in diameter (always more than 50% of sculpture), to a
mixture of grana, verrucae, bacula, and coni; occasionally either low coni or bacula
dominate the larger sculptural elements. Exine 0.4-0.8 um thick (exclusive of sculpture),
single layered. Equatorial diameter (100 specimens) ranges from 22 um to 62 um (median
about 40 um), 30% of specimens between 33 um and 43 um. A few abnormal monolete and dilete
(chevron) grains observed.
Remarks: Playford and Dettmann (1965) considered Concavisporites infirmus
Balme (equatorial diameter 35-46 um, mean 40 um) to be distinct from Trachysporites
asper Nilsson (size 38-41 um) and their specimens, since C. infirmus
possesses kyrtomes and a finely rugose sculpture. However, Balme's illustrations bear
remarkable resemblance to specimens of the present study, most of which have some degree
of kyrtome development. The illustrated specimen of T. asper (Nilsson, 1958, Pl.
2, fig. 3) also has raised arcuate folds flanking the laesurae. The finely rugose
sculpture in the illustrations of C. infirmus is not obvious (the sculpture
appears to consist of discrete grana and verrucae); a basal incipient reticulum could be
described as finely rugose, because closely spaced sculptural elements would be united to
give an apparent lineation pattern (S.E.M. photomicrograph, Pl. 2, fig. 2). The lack of
obvious distinction between C. infirmus, T. asper, and our specimens
favors uniting these forms under the earliest name, C. infirmus. This species
conforms to the generic requirements of Granulatisporites ibrahim emend. PotoniÕ
& Kremp, which has priority over Concavisporites Pflug and Trachysporites
Nilsson.
Granulatisporites infirmus grades into Converrucosisporites cameronii
(de Jersey) Playford & Dettmann in our samples, but for practical reasons the two
forms are separated by gross sculptural differences. Granulatisporites subgranulosus
(Couper, 1958) Fisher 1972 differs from G. infirmus by having a much thicker,
double-layered exine (l.5-3.0 um thick).
Affinity: Granulatisporites infirmus has been recovered by the first
author from sporangia of Clathropteris meniscoides Brongniart at Locality 8.
Associated spore masses in sporangial preparations contain C. cameronii. Some
dispersed spore preparations from the megafossil plant layers are dominated by C.
cameronii, while others are dominated by G. infirmus, although both forms
are invariably present in the same preparation. Specimens of Equisetites spp. are
the only other megafossils present in the plant bed. Probable roots and rhizomes of C.
meniscoides and Equisetites are preserved in situ. Only one species of Clathropteris
appears to be present, although it exhibits a wide range in frond site.
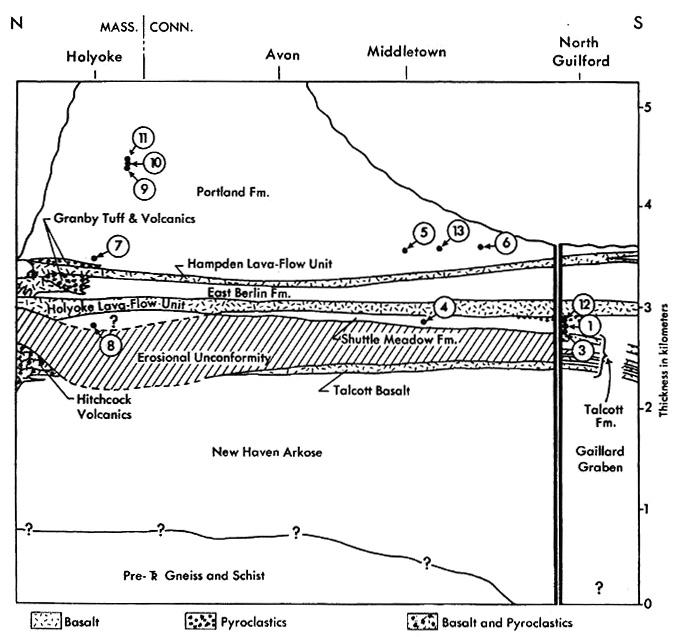
Localities: Portland Fm., 5, 11 (rare to scarce); Shuttle Meadow Fm., 1, 3, 4,
12 (rare to common); upper New Haven Arkose, 8 (abundant to dominant).
Genus Harrisispora Pocock 1970
Type species: Harrisispora mollis (Bolkhovitina 1956) Pocock 1970
Basionym: Adiantum mollis Bolkhovitina 1956
Remarks: See Pocock (1370, p. 38, 39) for generic description and generic
synonymy.
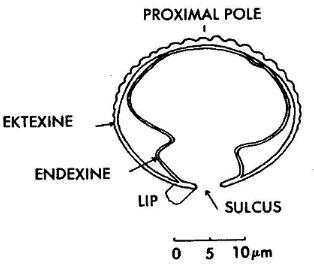
Harrisispora sp. 1
Plate 3, figure 3
Description: Miospores, trilete. Amb subtriangular with slightly convex sides.
Radius of laesura about 30% of diameter of spore; laesura occupies for nearly its entire
length a narrow zone or groove (1.6-3.0 um wide) of exinal thinning where ektexine and
endexine apparently separate. Both endexine and ektexine participate in forming the
laesura. Narrow, raised lips present on surface of endexine in groove. Adjacent to the
narrow zone of thinning, a band of exine (67 um wide) is inaapunctate and thickened on
each side of laesura. Exine double layered, psilate, about 1.4-1.6 um thick (exclusive of
proximal area of thickening); ektexine 1.0 um thick, endexine 0.4-0.6 um thick. Size (1
specimen): 80 um in diameter.
Affinity: Possibly ferns related distantly to Lygodium (Pocock, 1970).
Locality: Portland Fm., 11 (rare).
Genus Leptolepidites Couper 1953
Type species: Leptolepidites verrucatus Couper 1953
Leptolepidites cf. L. major Couper 1958
Plate 2, figures 11, 12
Remarks: Two specimens, measuring 46 um and 54 um, agree with L. major
in size, and in having verrucae frequently in contact. However, the verrucae of our
specimens are much smaller (1.0-2.5 um high; 1.0-3.0 um in diameter), agreeing in size and
in number around the edge with Leptolepidites bossus (Couper) Schulz. Our
specimens appear to be intermediate between L. major and L. bossus.
Locality: Portland Fm., 11 (rare).
Genus Murospora Somers 1952
Type species: Murospora kosankei Somers 1952
Murospora sp. 1
Plate 1, figures 9, 10
Description: Miospores, trilete. Amb subcircular to subtriangular in polar
view; equatorial outline of cingulum generally parallels that of central body. Laesura
extends almost to, sometimes reaching, equatorial margin of central body. Radial apices
occasionally protrude beyond general outline of central body, producing a bulge on the
cingulum. Laesura usually bordered by narrow, raised lips. Exine double layered, psilate.
Ektexine extends over whole surface, including cingulum. Endexine generally restricted to
central body, occasionally extending as lobes into, sometimes to outer margin of,
cingulum. Exine 0.9-1.0 um thick, ektexine about 0.4 um thick, endexine about 0.6 um
thick. Size range (7 specimens): 26-53 um in overall diameter (median 37 um); diameter of
central body 22-41 um; width of cingulum 3-7.0 um.
Locality: Portland Fm., 11 (rare).
Genus Perotriletes Couper 1953
Type species: Perotriletes granulatus Couper 1953
Perotriletes cf. P. pseudoreticulatus Couper 1953
Plate 2, figure 10
Description: Miospores radial, trilete. Amb subtriangular in polar view.
Laesura extends almost to equatorial margin, flanked by narrow, raised lips. Exine
(exclusive of perispore) about 0.6 um thick; both proximal and distal faces sculptured
with irregular, occasionally anastomosing, ridges, 1-2 um high. Perispore closely fitting,
densely covered with small grana, 0.1-0.4 um high; perispore attached only at laesura
margins. Overall site (2 specimens) 55-56 um, about 52-53 um in diameter, excluding
perispore.
Remarks: The presence of grana on the perispore might be sufficiently
different from the type description to warrant the creation of a new species. However,
because of the small number of specimens, a tentative assignment is made until further
information can be obtained. Our specimens differ significantly from other described
species of Perotriletes.
Locality: Portland Fm., 11 (rare).
Genus Todisporites Couper 1958
Type species: Todisporites major Couper 1958
Todisporites rotundiformis (Malyavkina 1949) Pocock 1970
Plate 2, figures 13-15
Cyclina pseudolimbata var. rotundiformis Malyavkina, 1949, p. 53, P1.
9, fig. 13.
Leiotriletes rotundiformis (Mal.) Bolkhovitina, 1956, p. 30, P1. 1, fig. 42b.
Todisporites minor Couper, 1958, p. 135, P1. 16, fig. 9, 10.
T. rotundiformis (Mal.) Pocock, 1970, p. 30, P1. 5, fig. 15.
Description: See Couper (1958, p. 135).
Remarks: Our specimens are very similar to T. minor, but have a
thinner exine (0.4-0.7 um) and a distribution curve skewed toward smaller size.
Measurement of 32 specimens from Locality 11 gives a size range of 29 um to 54 um (median
39 um, mean 40 um, mode 35 um). However, these differences are not significant. Pocock
(1970) gives a size range of 38 um to 46 um, Couper (1958) gives 32 um to 50 um (mode 45
um), Malyavkina (1949) gives 30 um to 40 um. Some of our larger specimens are
subtriangular in shape, but invariably these have major folds.
Affinity: Couper (1958) compared T. minor with the spores of Todites
princeps (Presl) Gothan, which are smaller (20-40 um) and have a mean (32 um) close
to our mode.
Localities: Portland Fm., 10, 11 (rare).
Genus Verrucosisporites Ibrahim 1933 emend.
PotoniÕ & Kremp 1954
Type species: Verrucosisporites verrucosus Ibrahim 1933
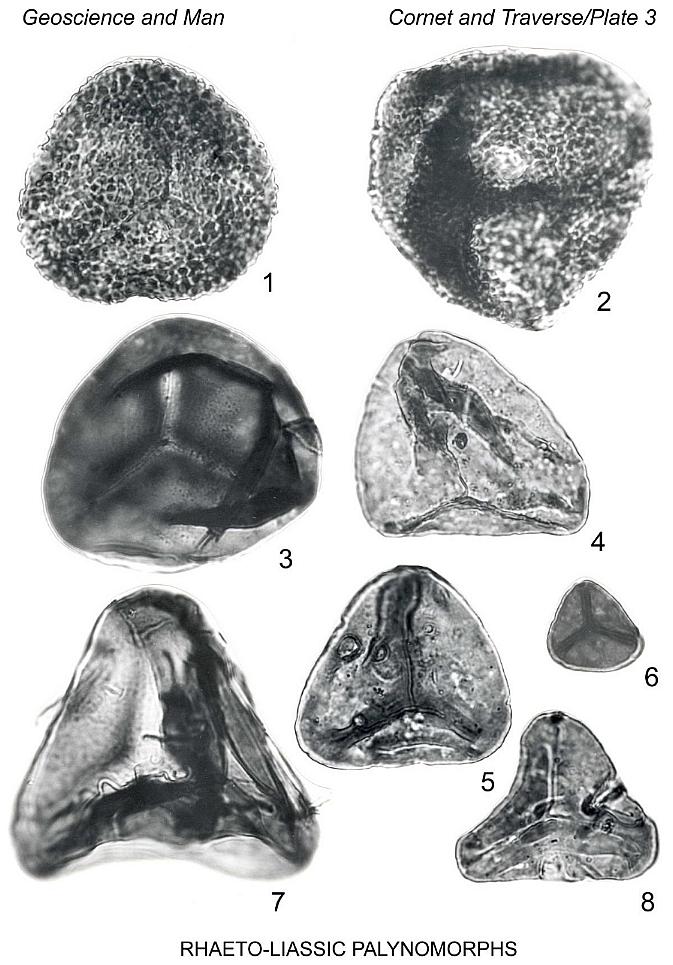
Verrucosisporites cheneyi n. sp.
Plate 3, figures 1, 2
Holotype: Pl. 3, fig. 1, size 73 um; Slide BS3, coordinates 40.5 x 122.7, ref.
56.2 x 126.2.
Description: Isospores, radial, trilete. Amb circular to subcircular, proximal
face less convex than distal side. Exine densely sculptured with verrucae and a few grana,
0.9-3.4 um high, 1.5-2.8 um wide. Verrucae sometimes in contact with one another,
occasionally fused into larger elements. Radius of laesurae variable, 26-78% of equatorial
diameter. Exine (exclusive of sculpture) approximately 0.8 um thick, single layered.
Equatorial diameter (41 specimens) 58-83 um (median 70 um), 90% of grains between 59 um
and 79 um.
Remarks: This species compares closely with the Permian Verrucosisporites
trisectus Balme & Hennelly 1956b, but differs somewhat in size range (V.
trisecatus: 65-135 um), and in the degree of suture development (laesurae of V. trisecatus
frequently extend onto distal surface of spore). The sculpture of both species is very
similar.
Species most prevalent in lake bed at Locality 1.
Species named after Jane Cheney, director emeritus of the Children's Museum of West
Hartford, Conn., in recognition of her efforts to preserve the natural history of the
Connecticut Valley.
Affinity: Osmundaceous fern?
Localities: Portland Fm., 6, 11, 13 (rare); Shuttle Meadow Fm., 1 (rare);
upper New Haven Arkose, 8 (rare).
Spore A
Plate 3, figure 7
Description: Miospore, trilete. Amb triangular in polar view, apices rounded.
Radius of laesura about 30% of diameter of spore. Laesura open, bordered by a kyrtome
consisting of narrow bands of exinal thickening. Distai side extended, equatorially
strongly concave, forming triradiate fold centered at distal pole. Exine double layered;
endexine 0.7 um thick; ektexine 1.0 um thick, loosely attached along triradiate fold.
Size: 98 um in diameter.
Locality: Portland Fm., 11 (rare).
Genus Pilasporites Balme & Hennelly 1956a
Type species: Pilasporites calculus Balme & Hennelly 1956a
Pilasporites allenii Batten 1968
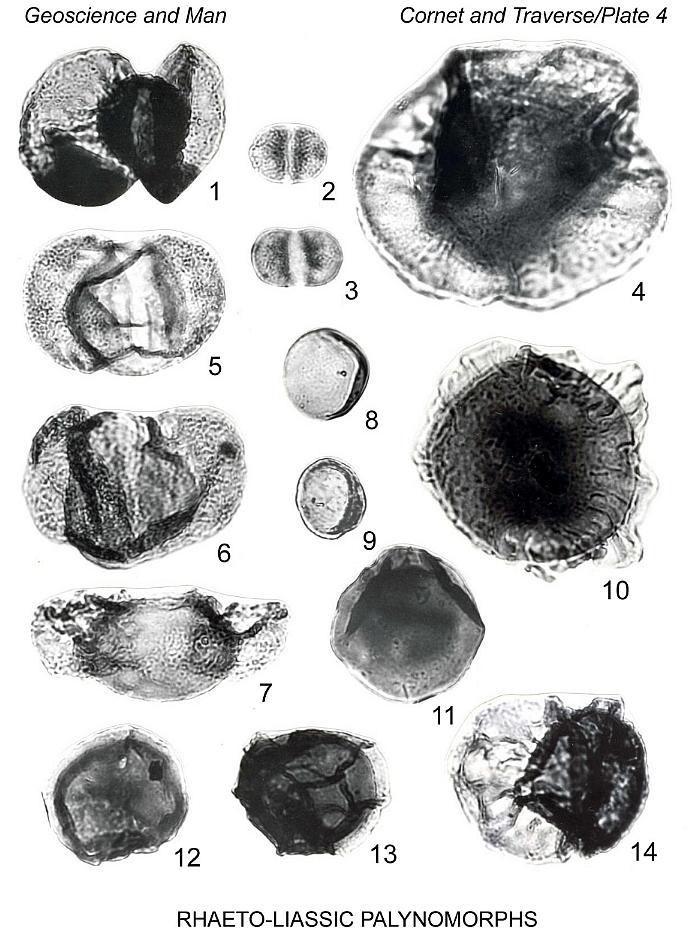
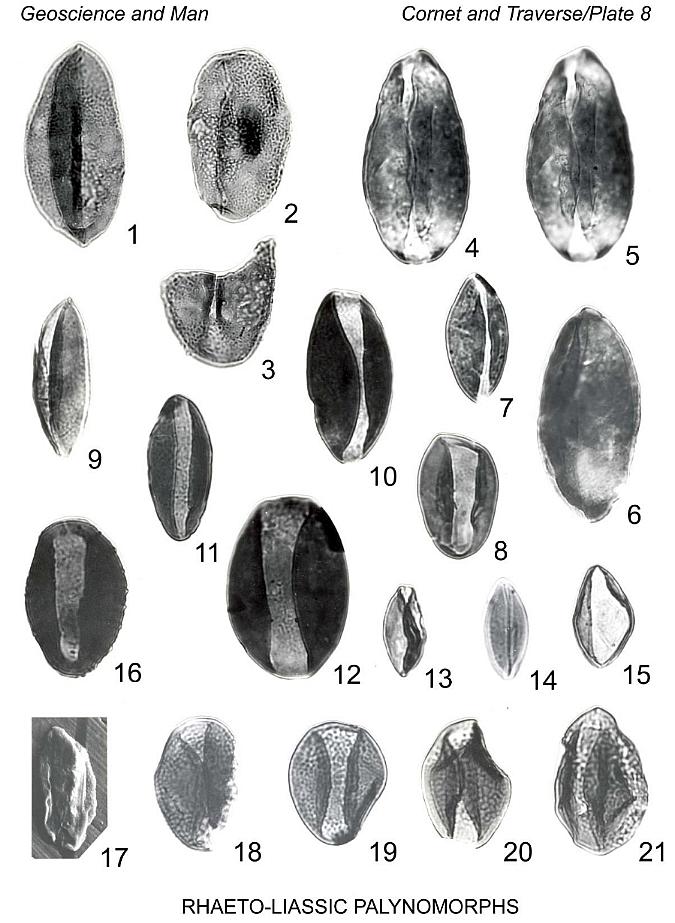
Plate 4, figure 11
Description: See Batten (1868, p. 638).
Remarks: Our specimens agree closely in all respects with P. allenii Batten
from the Lower Cretaceous of England, except in exine thickness, which is not a
significant difference for the creation of a new species. Measurement of 72 specimens from
Locality 8 gives a size range of 30 um to 54 um (median 39 um, mean 39 um), indicating a
slightly larger average size than that of the type description (mean 36.7 um; Batten,
1968). Exine of our specimens is 0.5-0.8 um thick as compared with 1.25-1.75 um given by
Batten. Slightly irregular distribution in the size-range curve of our specimens suggests
more than one source plant, as is also indicated by megafossils.
Affinity: Equisetites cf. E. sarranii Zeiller (abundant in
layers containing P. allenii), as well as possibly two other species of Equisetites,
present at Locality 8.
Localities: Portland Fm., 5, 2 1 (rare); Shuttle Meadow Fm., 1 (rare in lake
bed); upper New Haven Arkose, 8 (rare to scarce).
Genus Araucariacites Cookson 1947
Type species: Araucariacites australis Cookson 1947
Remarks: Cookson (1947), following the suggestions of Erdtman (1947), created
the "sporotype" names Granulonapites (Ý = form-generic name) and Araucariacites
(Ý = organ-generic name), as alternative names to be combined with the epithet,
australis. The 1972 I.C.B.N. restriction on the use of alternative names is retroactive
only to January 1, 1953. Consequently, we consider subsequent attempts to validate one of
the two generic names (Couper, 1953, Nilsson, 1958) unnecessary, since Cookson's monotypic
species was validly published. Her sporotypes and sporomorphs are handled normally in all
respects as genera and species, respectively. Our choice of the generic name, Araucariacites,
is based on the fact that Cookson does not accept the name Granulonapites in her
publication: wherever the generic concept is given a single name, that name is always Araucariacites.
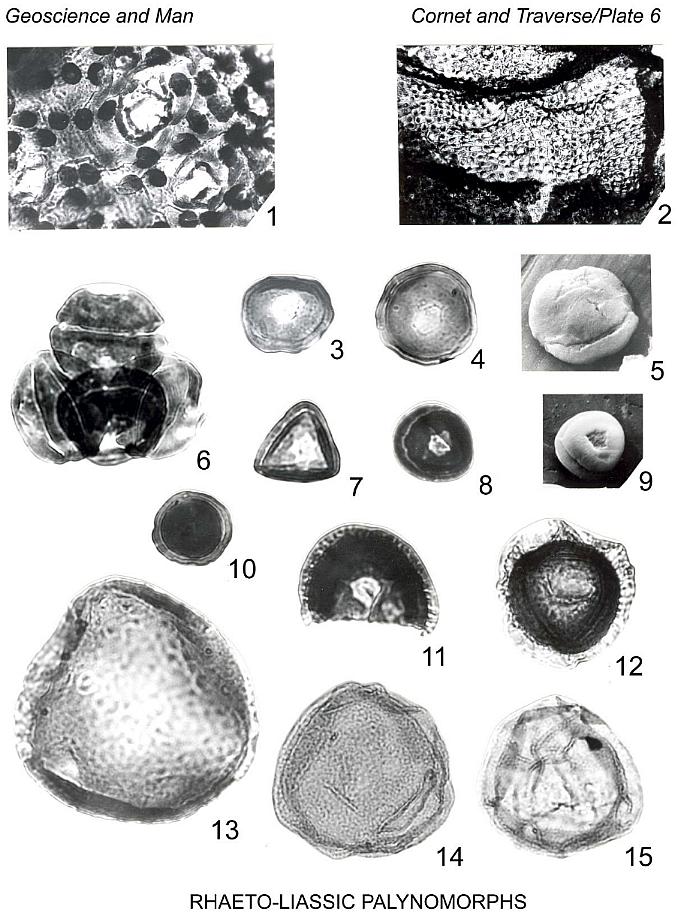
Araucariacites punctatus (Nilsson 1958)
comb. nov.
Plate 6, figures 14, 15
Granulonapites punctatus Nilsson, 1958, p. 70, Pl. 5, fig. 22.
Description: Pollen grains inaperturate, spherical to oval, frequently folded.
Exine 0.5-0.7 I~m thick, densely sculptured with small grana, 0.2 um or less in diameter.
Size range (50 specimens): 28-63 um (median 44 um) in maximum width or diameter, 90% of
specimens between 34.0 um and 56.8 um.
Remarks: This species agrees in all morphological respects with Araucariacites
australis Cookson, but has a smaller mean and smaller site distribution than
generally recognized for A. australis. Couper (1953) reported grains of similar
size under the name A. australis from Jurassic-lower Oligocene strata of New
Zealand. Nilsson (1318) described one specimen (upon which A. punctatus is based)
from the middle Keuper of Scania.
This species is generally much more common than A. australis in our samples.
Localities: Portland Fm., 5, 6, 7, 9, 10, 11, 13 (rare to scarce); Shuttle
Meadow Fm., 1, 12 (rare).
Araucariacites australis Cookson 1947
Plate 6, figure 13
Remarks: This species is distinguishable from A. punctatus (Nilsson) comb.
nov. by its larger size and usually darker color. Range in size for 26 specimens: 54-94 um
(median 73 um). Our specimens compare well in shape and in manner of folding to grains of
Brachyphyllum mamillare Brongniart as illustrated by Couper (1958, Pl. 27, fig. 1, 2).
Species is more frequently encountered in the Portland Fm. than in older strata. Some
specimens assigned to this species may be unsplit examples of A. fissus Reiser &
Williams.
Localities: Portland Fm., 5, 11 (rare); Shuttle Meadow Fm., 1 (rare).
Araucariacites fissus Reiser & Williams 1969
Plate 7, figures 6, 7
Description: See Reiser & Williams (1969, p. 17).
Remarks: The size distribution (17 specimens) indicates a larger average size
than that given by Reiser & Williams (1969), being 58-110 um (median 85 um) rather
than 34-95 um (?mean 57 um) for the Australian forms. The indicated Early Jurassic age for
A. fissus Reiser & Williams agrees well with the proposed age for the Portland
Fm. This species becomes more frequent in younger strata. The largest forms are more
common at Locality 5.
Discussion of relationship of A. fissus to Triangulopsis: Our
specimens (esp. P1. 7, fig. 7) from Locality 11 compare well in overall shape, manner of
splitting, and size range to Triangulopsis discoidalis D—ring 1961, recorded
from the Upper Jurassic of Germany and western Canada. However, T. discoidalis
apparently has a much thicker, double-layered exine, separable into an outer envelope and
an internal body (Pocock, 1970, p. 74). D—ring considered T. discoidalis to be
microplanktonic, while Pocock's study of a large population of this species indicated
morphological affinity with vascular plants. D—ring considered Zonalapollenites
to be synonymous with Triangulopsis, suggesting a similar basic structure. It is
possible that A. fissus is related to Triangulopsis discoidalis, lacking
only the development of a distinct endexine.
Localities: Portland Fm., 5, 10, and 11 (rare).
Genus Perinopollenites Couper 1958
Type species: Perinopollenites elatoides Couper 1958
Perinopollenites elatoides Couper 1958
Plate 4, figures 12, 13
Description: See Couper (1958, p. 152).
Remarks: One specimen from~locality 11 clearly shows a pore (Pl. 4, fig. 12).
Other specimens possess a highly folded "perine," which may simulate a crude
reticulum (Pl. 4, fig. 13). Specimens from Locality 5 generally have a slightly smaller
body, giving the appearance of a relatively larger "perine." Six specimens range
in maximum diameter from 34 um to 52 um; diameter of central body 28 um-43 um.
Localities: Portland Fm., 5, 11 (rare).
Genus Callialasporites Sukh Dev 1961
Type species: Callialasporites trilobatus (Balme 1957) Sukh Dev 1961 (designated by Sukh
Dev, 1961)
Basionym: Zonalapollenites trilobatus Balme 1957
Zonalapollenites (in part) Pflug in Thomson and Pflug, 1953, p. 66, 67.
Callialasporites Sukh Dev, 1961.
Applanopsis D—ring, 1961.
Triangulopsis (in part) Doring, 1961.
Pflugipollenites Pocock, 1962.
Applanopsipollenites Levet-Carette, 1964.
Description: See Sukh Dev, 1961, p. 48; Balme, 1957, p. 32.
Remarks: Sukh Dev (1961) and Pocock (1970) discuss the synonymy of Zonalapollenites
and Tsugaepollenites, and differences between these two genera and species now
placed in Callialasporites.
Callialasporites cf. C. dampieri (Balme 1957) Sukh Dev
1961
Plate 4, figure 10
Description: See Balme (1957, p. 32).
Remarks: Three specimens range in overall diameter from 60 um to 77 um; body
56 um-59 um in diameter. Sculpture of saccus is finely granulate. Endexine of body is
about 1.3 um thick. A slightly darkened area is present on the central part of the body.
One specimen has saccus folded back on body, resembling a similar condition found in C.
segmentatus (Balme) comb. nov.
Affinity: Gamerro (1965) found grains of C. dampieri and C.
trilobatus (Balme) Sukh Dev in male cones in organic connection with leaves of the
gymnosperm Apterocladus lanceolatus Archangelsky.
Locality: Portland Fm., 11 (rare).
Callialasporites segmentatus (Balme 1957) comb. nov.
Basionym: Zonalapollenites segmentatus Balme 1957, p. 33, Pl. 9, fig. 93, 94.
Remarks: A formal transfer is made of this species here, although no specimens
definitely referable to it were found.
Callialasporites trilobatus (Balme 1357) Sukh Dev 1961
Plate 4, figure 4
Description: See Balme (1957, p. 33).
Remarks: This species is larger than our specimens of Callialasporites
dampieri, 7 specimens measuring 78 um to 104 um in overall diameter; diameter of body
46 um-84 um. Exine of body is 0.8-1.1 um thick. Saccus is densely covered with small
grana, 0.2-0.8 um in diameter. The shape of the body ranges from oval to subtriangular,
with 3 lobes well developed on forms with subtriangular bodies. Forms with oval bodies
tend to possess sacci without indentations. Sukh Dev (1961) described a new species, Callialasporites
monoalasporus, that closely resembles our forms with oval bodies. More specimens are
needed to determine if a significant distinction exists between our forms resembling C.
monoalasporus Sukh Dev, and C. trilobatus. In addition, C. monoalasporus
may be conspecific with C. turbatus (Balme) Schulz 1967.
Affnity: Gamerro (1965) illustrates grains of C. trilobatus and
C. dampieri, as well as forms resembling C. monoalasporus, which he found
associated with Apterocladus lanceolatus Archangelsky. Reiser & Williams
(1963) found in their study forms of Callialasporites gradational between
different species of this genus, suggesting that many of them are derived from one source
plant.
Locality: Portland Fm., 11 (rare).
Genus Alisporites Daugherty 1941 emend. Nilsson
1958
Type species: Alisporites opii Daugherty 1941
Alisporites thomasii (Couper 1958) Nilsson 1958
Plate 4, figure 5-7
Pteruchipollenites thomasii Couper, 1958, p. 150, P1. 26, fig. 10, 11.
Alisporites thomasii (Couper) Nilsson, 1958, p. 83, P1. 8, fig. 1.
Description: See Couper (1958, p. 150).
Remarks: Our specimens compare closely with A. thomasii in overall
size and shape, as well as exine structure and sculpture. Twenty-one specimens range from
48 um to 77 um in overall breadth (median 69 um); breadth of body 20 um-46 um (median 36
um); length of saccus 36 um-57 um (median 40 um). Some specimens, not included in the
above measurements, have bodies compressed more or less obliquely with pendantly attached
sacci, producing forms suggestive of pollen of extant Pinus. More study is needed
to determine whether these unique forms should be included in A. thomasii.
Localities: Portland Fm., 5, 11 (rate).
Genus Pityopollenites Reissinger 1950 emend.
Jansonius 1962
Type species: Pityopollenites pallidus Reissinger 1950
Vitreisporites Leschik 1956 (imprint date, 1955).
Caytonipollenites Couper 1958.
Vitreisporites Leschik emend. Jansonius 1962.
Remarks: Pityopollenites was validly published by Reissinger in 1950,
and was monotypic at the time of publication. Vitreisporites, as originally
described by Leschik in 1956, required a faint laesura and a size of the central body
under 20 um. Jansonius (1962) removed these characteristics from the generic description,
considering them either not convincingly demonstrated or too restrictive. For further
discussion, see Dunay (1972).
Pityopollenites pallidus Reissinger 1950 emend. Nilsson 1958
Plate 4, figures 2, 3
Pityopollenites pallidus Reissinger, 1950, p. 105), P1. 15, fig. 1-5.
Vitreisporites signatus Leschik, 1956 (imprint date, 1955), p. 53, P1. 8, fig.
10.
Caytonipollenites pallidus (Reissinger) Nilsson, 1958, p. 78, P1. 7, fig. 12-14.
Description: See Nilsson (1958, p. 54).
Remarks: Twenty-one specimens of this study range fiom 22 um to 33 um in overall
breadth (median 27 um); breadth of body 7 um-15 um (median 27 um); length of saccus 14
um-22 um (median 17 um). Specimens from Locality 5 are generally smaller than those from
Locality 11.
Locatities: Portland Fm., 5, 11 (rare).
Genus Platysaccus Naumova 1939 emend. PotoniÕ
& Klaus 1954
Type species: Platysaccus papilionensis PotoniÕ & Klaus 1954
Remarks: Platysaccus was validly published by Naumova in 1939 (p.
355, fig. 1), although no type species was designated, which is not required by the
I.C.B.N. before 1358. PotoniÕ and Klaus (1954) designated their new species, P.
papilionensis, as type, and emended the genus.
Platysaccus cf. P. lopsiensis (Malyavkina 1961) Pocock
1970
Plate 4, figure 1
Podocarpus lopsiensis Malyavkina in Samoilovich et al., 1961, p. 130, Pl. 35,
fig. 9a,b.
Platysaccsus lopsiensis (Mal.) Pocock, 1970, p. 85, Pl. 18, fig. 6, 7.
Description: See Pocock (1370, p. 85).
Remarks: Two specimens compare closely with P. lopsiensis, measuring 56 um to
59 um in overall breadth (grains not completely expanded); breadth of body 26 um-27 um ;
length of saccus 26 um-46 um.
Locality: Portland Fm., 11 (rare).
Genus Podocarpidites Cookson 1947
Type species: Podocarpidites elliptica Cookson 1947 (designated by
Potonie', 1958)
Remarks: This generic name was validly published by Cookson (1947) under
circumstances similar to those described above under Araucariacites Cookson.
Podocarpidites sp. 1
Plate 4, figure 14
Description: Pollen grains, bisaccate. Body of grain generally longer than
broad; sacci strongly distally pendant, rarely expanded, usually highly folded. Distal
surface traversed by an elongate sulcus, about 20 um long, flanked by basal attachment of
sacci. When expanded, saccus measures about 36 um in length, 50 um in breadth. Proximal
surface of body covered by a closely adhering cappa, 1.4-2.6 um thick, tightly folded to
form a verrucose and rugose surface. Exine of body about 0.9 um thick. Sacci possess fine
reticulum with lumina 0.2-0.3 um in diameter. Size of body (5 specimens): 40-46 um long;
31-34 um broad. Sacci generally about 30 um in diameter (folded). Overall breadth
approximately 50-60 um, but highly dependent on shape of sacci.
Remarks: Bisaccates are generally rare in the Shuttle Meadow Fm. However, they
increase slightly in abundance in the lake bed and underclay at Locality 1. A similar
florule from Licking Run near Midland, Va. (see Discussion), contains as much as 15%
bisaccates in certain lacustrine horizons. Most bisaccates from floras dominated by Corollina
meyeriana compare best with Podocarpidites and Pityosporites,
although examples of Alisporites have been found. Generally poor preservation of
most of the Shuttle Meadow bisaccates prevents description of several other types present.
Localities: Shuttle Meadow Fm., 1, 12 (rare); upper New Haven Arkose, 8?
Circulina-Corollina-Classopollis
Nomenclatural problem: Circulina and Corollina were validly
published as generic names by Malyavkina (1949), even though her generic and specific
descriptions have been regarded as vague and inadequate for validation (Pocock and
Jansonius, 1961; Reyre, 1969; Traverse, Ames and Spackman [CCFSP, 1974, in press: 38-103,
38-128]). Classopollis is generally regarded as the acceptable generic name, but
was originally described as tricolpate by Pflug (1953). Subsequent emendation of Classopollis
by Pocock and Jansonius (1961) and Couper (1958) largely corrected the description of this
pollen type, but Chaloner (1962) and Pettitt and Chaloner (1964) provided the most
accurate description, clarifying the structure of the exine with electron micrographs of
well-preserved grains. Klaus (1960) emended the description of Circulina and Corollina,
providing Hilfstypus -- better called neotype, which is admissible under 1372
I.C.B.N., Guide for Determination of Types, p. 76, for establishing taxonomic validity.
However, Klaus chose to distinguish Circulina from Corollina, making Classopollis
a junior synonym of Corollina.
S. Samoilovich (Institute of Petroleum, Leningrad, USSR; pers. comm. to A. Traverse,
1973) has indicated that Malyavkina's type material was not preserved. She provided
photographs (Pl. 4, fig. 9; Pl. 5, fig. 10) of "neotypes" (chosen by Zauer,
supplied to Klaus, and chosen by him as Hilfstypus) for species of Circulina
and Corollina, derived fiom the same Lower Jurassic stratigraphic horizon as
studied by Malyavkina (from the same locality, a borehole 80 km to the west of the
settlement of Sagiz). Even though the assignment of these "neotypes" by Zauer is
subjective, it is perhaps the only way to obtain stability of nomenclature and to avoid
personal preference for the name Classopollis.
It is interesting to note that the "neotype" of Corollina compacta
Malyavkina (Pl. 5, fig. 10) compares well with Classopollis in exinal structure,
even though a well-developed distal pseudopore is not present, and equatorial striations
are questionably present. Thus Klaus was apparently correct in making Classopollis
synonymous with Corollina rather than with Circulina, It is further
significant that the "neotype" of Circulina simplex Mal. closely
resembles Malyavkina's illustrations, lacks a well-developed ring tenuitas (rimula) and
distal pseudopore, and resembles specimens of Spheripollenites Couper and
endoexinal bodies of Classopollis more than it does Circulina meyeriana
(Mal.) Klaus, raising the question of the correctness of Klaus's emendation of Circulina.
Reyre (1969) considers Circulina Mal. emend. Klaus as "an exceptional case within
the genus Classopollis, corresponding to the coincidence of the limiting cases of
the two characters, equatorial thickening and ornamentation of the intrastructure."
He describes two species, Classopollis quezeli and C. kieseri, with a
typical Classopollis intrastructure, but without equatorial striations or with
only vaguely defined striations (pseudostriations). These two species closely resemble
grains that have been erroneously assigned to Corollina meyeriana (Klaus)
Venkatachala & GoÓzÃn (Schulz, 1967, P1. 22, fig. 6) or Circulina meyeriana
Klaus (Geiger and Hopping, 1968, P1. 4, fig. 32). In fact this confusion suggests that
there is really no clear generic distinction between grains with a massive intrastructure
and no equatorial thickening or striations, and grains with a structured exine, equatorial
thickening, and striations.
Consequently the only valid genetic characteristics are those that Circulina
emend. Klaus and Classopollis have in common: frequent occurrence as tetrads,
spherical shape, size range (18-60 um), proximal trilete mark, distal pseudopore, and
subequatorial furrow or rimula. Since the intrastructure of the exine ranges from massive
through various types (Reyre, 1969), equatorial striations vary in number and
completeness, particularly in some species, for example, Classopollis torosus
(Reissinger) Balme and C. kieseri Reyre, and because the amount of equatorial
thickening is variable, even in a single species, these characteristics are best reserved
for specific distinction.
It is therefore clear that Circulina (as emended by Klaus) and Corollina
should be combined into a single genus, with Classopollis as a junior synonym. Circulina
Malyavkina may not be identical with Circulina Mal. emend. Klaus (the status of
which is also confused). Corollina was monotypic when published, whereas Circulina
was not. Thus, Corollina does not present problems of generic typification and is
therefore chosen to embody all of the common generic characters.
Genus Corollina Malyavkina, 1949 emend.
Corollina Malyavkina 1949.
Circulina Malyavkina (in part), 1949.
Classopollis Pflug 1953.
Classopollis Pflug emend. Couper 1958.
Corollina Mal. emend. Klaus 1960.
Circulina Mel. emend. Klaus 1960.
Classopollis Pflug emend. Pocock & Jansonius 1962.
Monilapollis Chang 1963.
Pagiophyllumpollenites Chang 1963.
Classopollis Pflug emend. Reyre 1969. [Also the species listed below referred to
"Pollenites" by Thiergart and by Reissinger.]
Generic type: Corollina compacta Malyavkina, 1949, p. 124, Pl. 46,
fig. 10. (Liassic).
Holotype: Malyavkina, 1949, P1. 46, fig. 10, 11.
Neotype: Plate 5, fig. 10. Designated by Zauer, photograph supplied by
Samoilovich, 1973.
Stratigraphic position and locality of neotype: Borehole Kozhegali K36, 466.8
m-471.9 m, Lower Jurassic, 80 km west of Sagiz near Caspian Sea.
Species assigned to genus: Circulina funifera Malyavkina, 1949, p.
123, P1. 46, fig. 6; Bennettiteae-Pollenites reclusus
Thiergart, 1949, p. 11, P1. 2, fig. 14, 15, P1. 3, fig. 6; Pollenites torosus
Reissinger, 1950, p. 115, P1. 14, fig. 20.
Emended generic description: Pollen grains distally pseudoporate, proximally
trilete, spherical to ovoid, or compressed, acorn shaped; occur as tetrads or isolated
grains; exine two layered, with distinct endexine and ektexine. Endexine comprises a
spherical, internally separate body, which frequently may be poorly developed and
difficult to distinguish, or well developed, sometimes occurring as separate bodies from
rupnued grains. Ektexine (modified from Reyre, 1969) composition variable on different
parts of the grain; composed of inner structured layer, comprising inuastructure, and
outer tegillum, which is continuous over entire grain and covered with outer sculpture
uniformly distributed over whole surface. Intrastructure massive, alveolate, punctate,
vermiculate or pseudovermiculate, reticulate or pseudoreticulate, but can be
heterogeneously organized: reduced, or thickened, in parts of the grain; reduced at distal
pole (pseudopore) or only along circular line surrounding it, reduced along a
subequatorial furrow (ring tenuitas or rimula) on distal side of equator, and sometimes
reduced at proximal pole (triangular area). Intrastructure generally thidcened in
equatorial zone on proximal side of furrow or rimula, but never thinner than ektexine
proximai to equatorial zone or belt; intrastructurai elements within this equatorial zone
may or may not be organized into more or less continuous circumequatorial striations; they
are absent if intrastructure of this zone is massive.
Remarks: Endexinal body may or may not possess a poorly developed trilete
mark; it infrequently possesses a distal area of thinning or tenuitas. The neotype of Circulina
simplex Malyavkina (our P1. 4, fig. 9) and illustrations by Malyavkina (Pl. 46, fig.
7-3) strongly resemble separated endexinal bodies of Corollina, and therefore do
not possess sufficient characteristics to place C. simplex in Corollina.
Circulina funifera Mal. (Pl. 46, fig. 6), on the other hand, appears to possess a
subequatorial furrow (border sharply differentiated from the body, narrow, double), and
exinal intrastructure (border covered with fine, radially directed, straight lines), which
possibly place it in Corollina.
Corollina torosus (Reissinger 1950) Klaus 1960 emend.
Plate 5, figures 2-3, 14
Pollenites torosus Reissinger, 1950, p. 115, Pl. 14, fig. 20.
Classopollis classoides Pflug, 1953, Fig. 4, J-M, P1. 16, fig. 20-25, 229-37;
emend. Pocock & Jansonius, 1961, p. 443, P1. 1, fig. 1-3.
Classopollis cf. C. torosus (Reissinger) Balme, 1957, p. 37, Pl. 11,
fig. 114-119.
C. torosus (Reissinger) Couper, 1958, p. 156, Pl. 28, fig. 2-7; Nilsson, 1958, p.
74, Pl. 7, fig. 68.
C. torosus (Reissinger) Balme; Chaloner, 1962, p. 13, fig. 2, Pl. 11, fig. 1, 2;
emend. Burger, 1965, p. 65, P1. 1, fig. 1, 2.
Monilapollis hsui Chang, 1963, p. 435, 439, P1. 1, fig. la-lc, 2a-2c.
"Classopollis torosus" Pettitt & Chaloner, 1964, P1. 1, fig. 1-5.
Emended description: Isolated pollen grains or occurring as atrads; spherical
to ovoid, or acorn-shaped; diameter in equatorial view (100 grains): 18-41 um (median 32
um), 90% of grains between 24 and 37 um (Couper, 1958, notes range 24-46 um, mode 32 um).
Diameter in polar view 24-37 um (diameter of subequatorial furrow 83-88% of overall
diameter). Distal pseudopore variable in size, even for grains of same size: 4.3-8.5 um in
diameter, frequently distorted and widened in equatorial view. Proximal triangular area of
thinning, which sometimes contains trilete mark, 6.8-13.6 um high. Exine two layered;
ektexine divisible into outer tegillum, continuous over whole surface of grain, and inner
structured layer, which is thickened in equatorial region to form a band or belt. Endexine
variably distinct to indistinct, spherical, frequently scabrate, occasionally with a small
proximal trilete mark. Structure of equatorial band ranges from pseudoreticulate, to
vaguely striate with aligned intrastructural columellae (pseudostriations), to distinctly
striate, with columellae fused to form more or less parallel bands, the number of
striations frequently varying from one part of band to another. Sometimes combinations of
grains with all degrees of striation development occur in same tetrad. When striations are
well developed, maximum number per grain ranges from 6 to 10. Width of equatorial band,
which frequently ranges from 5 um to 13 um at its maximum, usually decreases in one area
to 4-10 um, and striations decrease in number to 4 or 5 through anastomosis or
disintegration of more proximal striations into discrete intrastructural elements. Exine
thickness varies with grain size, and presumably also with preservation; it is thickest in
equatorial band: 1.3-2.4 um (Couper, 1958, records 3.0 um), decreasing gradually toward
proximal pole: for example, 2.l um down to 0.9 um; thickness on distal side of equaterial
furrow (range 0.7-1.7um) generally slightly less than that proximal to equatorial band.
Intrastructure of ektexine mostly pseudoreticulate, with positive elements largest on
proximal side of equatorial furrow, particularly if equatorial band is pseudoreticulate;
pseudoreticulum on distal side of grain usually finer in pattern, occasionally becoming
punctate. Sculpture of tegillum variable (perhaps owing to preservation); light microscope
indicates roughened surface; S.E.M. indicates microscabrate sculpture, mixed with
microverrucae and microconi (Pl. 5, fig. 14; see also Pettitt and Chaloner, 1964, p. 614);
sculpture distinctly less well developed than in species figured by Reyre (1969), perhaps
less well developed than in forms described under Classopollis torosus
(Reissinger) Couper emend. Burger 1965.
Remarks: The emended description of Classopollis classoides Pflug by
Pocock and Jansonius (1961) is not adequate and conflicts with our present knowledge of
the exine structure of this genus. We find no distinction between C. classoides
and C. torosus, these species agreeing in exinal structure and variation in
striation development (Pocock and Jansonius, 1961, Pl. 1, fig. 1-9). The size ranges
overlap widely, but the mean equatorial width for C. classoides (29 um) is
somewhat smaller than that of C. torosus (32 um). However, Harris (1957) noted a
still smaller mean in his samples (25 um) for grains placed by Pettitt and Chaloner (1964)
in C. torosus. These differences may result in part from preservational states
and preparation techniques. The mean diameter for C. torosus from Locality 11 is
32 um, while it is 25 um from Locality 5. Table 1 gives data for relative abundance of C.
torosus that suggest the existence of two separate "Circumpolles"
populations, intermixing at Locality 5. The morphology of these two populations does not
permit their distinction in a mixed sample, as the ranges of variation widely overlap.
Therefore, only one form-species is described, although two different source plants may be
indicated.
Affinity: Hirmerella muensteri (Schenk) Jung 1968, possibly Pagiophyllum
connivens Kendall 1948.
Localities: Portland Fm. 5, 6, 7, 9, 10, 11 (dominant); Shuttle Meadow Fm. 1,
3, 4, 12 (rare to scarce); upper New Haven Arkose, 8 (scarce).
Corollina itunensis (Pocock 1962) comb. nov.
Plate 5, figure 1
Classopollis itunensis Pocock, 1962, p. 71, Pl. 11, fig. 176, 177; P1. 12,
fig. 178 (Basionym).
Description: See Pocock (1970, p. 104).
Table 1. Approximate percentages of major Corollina species in Hartford Basin
(Based on Corollina counts only. Most of palynoflorules counted were at least 90%
Corollina.)
Localities arranged in stratigraphic order, youngest at right: N.H.A. = New Haven
Arkose; S.M.F. = Shuttle Meadow Formation; P.F. = Portland Formation; + = present, but
less than 1%.